
")
Figure 1. Geographical distribution of brown root rot disease (BRRD) worldwide and in Taiwan. Data sources: (1) Invasive Species Compendium, CAB International. www.cabi.org/isc. (2) Taiwania, International Journal of Biodiversity [1].
Brown root rot disease (BRRD) caused by Phellinus noxius (Corner) Cunningham, destroys the root interior and stem tissue of healthy trees. BRRD can be transmitted through root contact, contaminated soil, groundwater, and even air. BRRD has now been reported to be a crucial cause of tree decline and mortality throughout the Americas, Africa, Asia, and Oceania (Figure 1). In Taiwan, BRRD has become a severe threat that has been destroying the agriculture, landscape, and forest trees for the last 20 years [1]. Previous reports indicated that more than 200 agricultural and forest plant species are hosts of P. noxius. Half of the hosts were reported for the first time from Taiwan [2]. The extremely diverse host range and wide geographical distribution of P. noxius has become a severe threat to forest plantations, fruit orchards, and even urban environments.
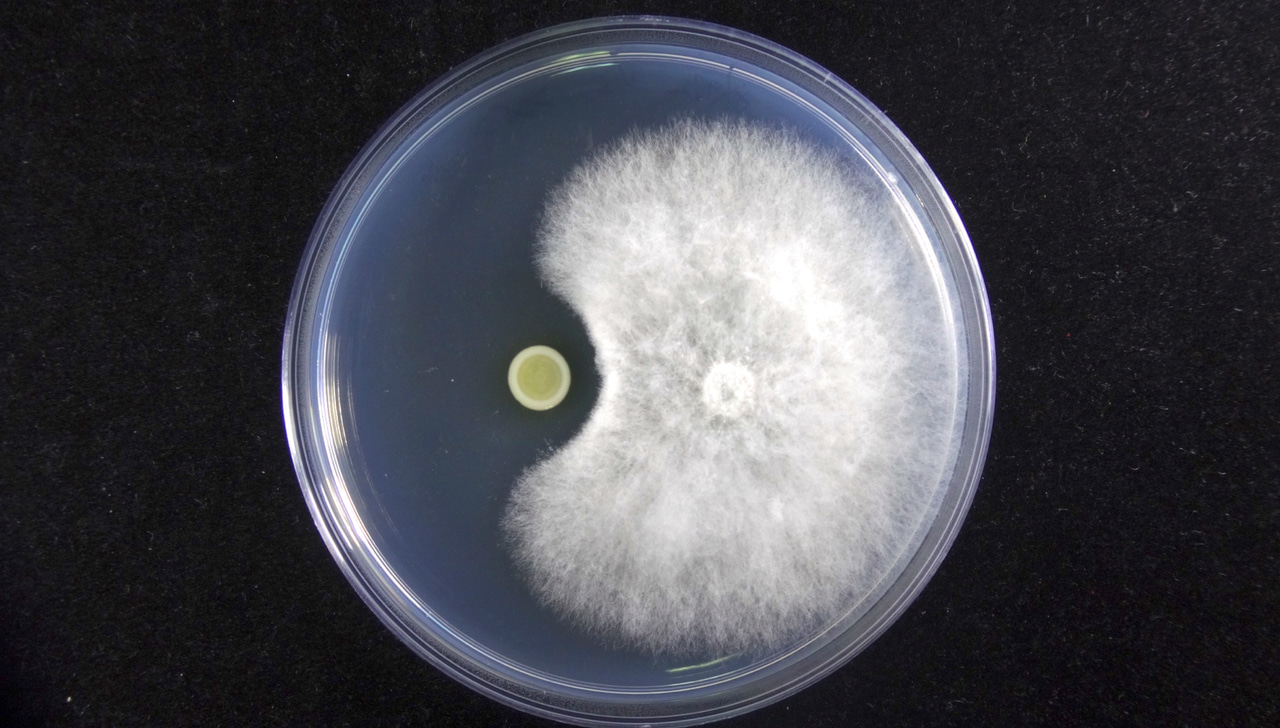
Several treatments have been applied to deal with BRRD, such as fungicide application, fumigation, and the removal of diseased plants. In Taiwan, chemical fungicides and fumigation are the major approaches used to combat BRRD [3, 4]. However, because of the environmental and health issues associated with the use of fungicides in agriculture, biological control of plant pathogens has come to be considered a more eco-friendly approach. Bacillus, Trichoderma, Streptomyces, and Pseudomonas species have been reported to be potential biocontrol agents against P. noxius [5-7], and produce many antifungal metabolites. Among these metabolites, pyochelin and pyoverdine, two well-known siderophores with antifungal activity, are commonly produced by Pseudomonas and Burkholderia species [8-10].
Recently, Dr. Yu-Liang Yang’s group reported that P. noxius effectively eliminates the antifungal activity of pyochelin by transforming pyochelin into pyochelin-GA. Meanwhile, dehydroergosterol peroxide is accumulated in fungal cells in response to pyochelin (Figure 2). The metabolic interactions between Burkholderia cenocepacia strain 869T2 and P. noxius were observed in situ using imaging mass spectrometry. Yang’s group further confirmed that the iron-chelating ability of pyochelin-GA is significantly decreased, which might influence the extracellular capture of iron using pyochelin by bacteria. The in silicon analysis revealed that the glycolic ester moiety of pyochelin-GA might affect the iron-chelating ability and decrease the pyochelin-GA binding ability to FptA, the outer membrane receptor of pyochelin-iron complex in Pseudomonas species. In addition, P. noxius was found to be able to transform ent-pyochelin to ent-pyochelin-GA, which demonstrates how P. noxius competes with the pyochelin and ent-pyochelin producer bacteria in the soil environment.
Overall, the study illustrates how fungal phytopathogens inactivate the antifungal and iron-chelating ability of bacterial siderophores. When the functions of bacterial siderophores were eliminated, the efficacy of bacterial biocontrol agents was severely weakened. Detailed information is available in the article entitled “Specific inactivation of an antifungal bacterial siderophore by a fungal plant pathogen” published by ISME Journal: https://doi.org/10.1038/s41396-020-00871-0.

Figure 2. Imaging mass spectrometry revealed the metabolic interaction in the battle between bacteria and fungi. The red color represents the distribution of pyochelin, and the green represents the distribution of pyochelin-GA. Ergosterol and dehydroergosterol are the main components of the fungal cell membrane.
References:
- Hsiao, W.W., T.H. Hung, and E.J. Sun., Newly discovered basidiocarps of Phellinus noxius on 33 tree species with brown root rot disease in Taiwan and the basidiospore variations in growth rate. Taiwania, 2019. 64(3): p. 263-268.
- Ann, P.J., T.T. Chang, and W.H. Ko, Phellinus noxius Brown Root Rot of Fruit and Ornamental Trees in Taiwan. Plant Dis, 2002. 86(8): p. 820-826.
- Tsai, J.N., P.J. Ann, and W. Hsieh, Evaluation of fungicides for suppression of three major wood-decay fungi Phellinus noxius, Rosellinia necatrix and Ganoderma australe in Taiwan. Plant Pathology Bulletin, 2005. 14: p. 115-124.
- Fu, C.H., et al., Evaluation of dazomet as fumigant for the control of brown root rot disease. Pest Manag Sci, 2012. 68(7): p. 959-62.
- Huang, H., et al., The effect of phenazine-1-carboxylic acid on the morphological, physiological, and molecular characteristics of Phellinus noxius. Molecules, 2016. 21(5): p. 613.
- Chou, H., et al., In Vitro and in Planta Evaluation of Trichoderma asperellum TA as a Biocontrol Agent Against Phellinus noxius, the Cause of Brown Root Rot Disease of Trees. Plant Disease, 2019. 103(11): p. 2733-2741.
- Leung, K.-T., et al., Brown Root Rot Disease of Phyllanthus myrtifolius: The Causal Agent and Two Potential Biological Control Agents. Plant Disease, 2020. 104(11): p. 3043-3053.
- Cox, C.D., et al., Pyochelin: novel structure of an iron-chelating growth promoter for Pseudomonas aeruginosa. Proceedings of the National Academy of Sciences, 1981. 78(7): p. 4256.
- Darling, P., et al., Siderophore production by cystic fibrosis isolates of Burkholderia cepacia. Infection and immunity, 1998. 66(2): p. 874-877.
- Briard, B., et al., Interactions between Aspergillus fumigatus and Pulmonary Bacteria: Current State of the Field, New Data, and Future Perspective. Journal of fungi (Basel, Switzerland), 2019. 5(2): p. 48.




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in